|
|
 2
mediates nuclear
import of
a mRNA binding protein 2
/ Ran / GTP hydrolysis / repeat nucleoporins)
2
mediates nuclear
import of
a mRNA binding protein 2
/ Ran / GTP hydrolysis / repeat nucleoporins)
Laboratory of Cell Biology, Howard Hughes Medical Institute, The Rockefeller University, 1230 York Avenue, New York, NY 10021
Contributed by Günter Blobel, March 6, 1997
 ABSTRACTINTRODUCTIONMATERIALS
AND METHODSRESULTSDISCUSSIONFOOTNOTESACKNOWLEDGEMENTSABBREVIATIONSREFERENCES
ABSTRACTINTRODUCTIONMATERIALS
AND METHODSRESULTSDISCUSSIONFOOTNOTESACKNOWLEDGEMENTSABBREVIATIONSREFERENCESABSTRACT
We have cloned and sequenced cDNA for human karyopherin INTRODUCTION Import of
proteins containing a nuclear localization
sequence (NLS) into digitonin-permeabilized cells is mediated by soluble
transport factors. A heterodimer, termed karyopherin
or importin, recognizes
the NLS protein in the cytoplasm via its Studies in yeast have revealed the existence so far of
three proteins that are both structurally and functionally related
to Kap95p (ref. 19; M. P. Rout, G.B., and J. D.
Aitchison, unpublished data) and therefore have been classified
as members of the yeast A detailed characterization of the
yeast Kap104p was recently reported (19). A cytosolic
complex could be isolated that contained Kap104p and two abundant
nuclear mRNA
binding proteins, Nab2p and Nab4p, with Nab4p being the likely
homolog of the vertebrate nuclear
mRNA-binding protein A1. This complex
did not contain karyopherin Here we describe the further functional characterization of
Kap104p's human homolog that we have termed karyopherin MATERIALS AND METHODS The GenBank cDNA clone 224297 (accession number R54232
[GenBank])
containing a sequence that was similar to yeast karyopherin To obtain a full-length cDNA for karyopherin To obtain a cDNA for the fusion protein GST-human nuclear
mRNA binding protein A1, the cDNA clone
81773 (Genome Systems) coding for full-length A1 was amplified
by the PCR primers 5 Recombinant human karyopherin Peptides corresponding to the NLS of
simian virus 40 T antigen (24) and the M9 sequence of
the human nuclearmRNA binding
protein A1 (21) were synthesized with an N-terminal
Cys for chemical coupling reactions.
The assay was performed essentially as described (16).
GST-A1 (1.5 ľg) was immobilized on 10 ľl of
glutathione beads, and 1 ľg of
karyopherin Proteins of rat liver nuclear
envelopes (29) or E. coli lysates expressing
Nup98 fragments (28) were separated by SDS/PAGE
and transferred to nitrocellulose. The blot was blocked for 1
h at room temperature in 5% milk/0.2% Tween 20 in transport buffer
[20 mM HepesˇKOH, pH 7.3/110 mM KOAc/2 mM Mg(OAc)2/1 mM
EGTA/2 mM DTT] and then incubated for 1 h at room temperature
in the same buffer containing Import assays were performed on
digitonin-permeabilized HeLa cells essentially as described (24),
except that GTP was added at 1 mM. Where specified, GTP was
substituted by 2 mM guanylyl imidodiphosphate.
When included, proteins were added to give the following final concentrations
per assay: 0.5 ľg of karyopherin RESULTS We searched the Expressed Sequence Tag database and found several sequences
that were similar to yeast Kap104p. Using an antisense oligonucleotides
derived from one of these sequences
(see
Materials and Methods), we screened a human liver
cDNA library and obtained several clones. From overlapping cDNA
clones we determined the DNA sequence and obtained a complete,
cDNA-deduced amino acid sequence for the protein that we termed
human karyopherin
We assembled a cDNA coding for full-length
Karyopherin
The binding site for karyopherin
To assay for docking to the nuclear
rim, digitonin-permeabilized cells were incubated on ice in transport buffer
containing fluorescently labeled GST-A1, with or without recombinant
To assay for import, digitonin-permeabilized
cells were incubated at room temperature with fluorescently labeled GST-A1,
an ATP-generating system, GTP, in the absence or presence of
various concentrations of Our finding (see above) that DISCUSSION From this and other recent studies (refs. 19, 22,
30; M. P. Rout, G.B., and J. D. Aitchison, unpublished
data) it appears that the importof
nuclear proteins occurs by at least
three different pathways in mammalian cells (or yeast). Proteins
are directed into these pathways by distinct NLSs and by cognate
NLS recognition and docking factors of
the karyopherin family. As a more general
nomenclature we suggest the generic terms karyopherin In this paper we have focused on the characterization of
a mammalian
karyopherin Recombinant GTP hydrolysis is required for In summary, karyopherin2,
also known as transportin. In a solution binding assay, recombinant2
bound directly to recombinant nuclear
mRNA-binding protein A1.
Binding was inhibited by a peptide representing A1's previously characterized
M9 nuclear localization sequence (NLS),
but not by a peptide representing a classical NLS. As previously
shown for karyopherin1,
karyopherin2
bound to several nucleoporins containing characteristic peptide
repeat motifs. In a solution binding assay, both1
and 2 competed with each
other for binding to immobilized repeat nucleoporin Nup98. In
digitonin-permeabilized cells, 2
was able to dock A1 at the nuclear
rim and to import it into
the nucleoplasm. At low concentrations of2,
there was no stimulation ofimport
by the exogenous addition of the GTPase
Ran. However, at higher concentrations of2
there was marked stimulation of
import by Ran.
Import
was inhibited by the nonhydrolyzable GTP analog guanylyl imidodiphosphate
by a Ran mutant that is unable to hydrolyze GTP and also by
wheat germ agglutinin. Consistent with the solution binding
results, karyopherin2
inhibited karyopherin /1-mediated
import of
a classical NLS containing substrate and, vice versa, 1
inhibited 2-mediated
import
of A1 substrate, suggesting that
the two import pathways merge at the
level of docking of1
and 2 to repeat nucleoporins.
/1-mediated
import of
a classical NLS containing substrate and, vice versa, 1
inhibited 2-mediated
import
of A1 substrate, suggesting that
the two import pathways merge at the
level of docking of1
and 2 to repeat nucleoporins.
subunit and, via its
subunit, docks the complex to a subset of
peptide repeat containing nucleoporins (1-10).
The GTPase Ran (11, 12) and
a Ran interacting protein, termed p10 (or NTF2) (13,
14), then mediates
release and GTP-hydrolysis-dependent transport of
the NLS protein and
karyopherin
into the nucleus with karyopherin
staying behind at the nuclear pore
complex (10, 15-17). Homologs
of these transport factors also
have been identified in yeast and recombinant Kap60p/Srp1p (karyopherin)
and Kap95p (karyopherin)
can substitute for their mammalian homologs in docking NLS protein
to the
nuclear rim of
digitonin-permeabilized mammalian cells (18).
karyopherin family. All four yeast
karyopherins [Kap95p, Kap104p,
Pse1p (20), and Kap123p] have been shown to serve
as transport factors for nuclear
protein import (ref. 19;
M. P. Rout, G.B., and J. D. Aitchison, unpublished data).
.
Thus, unlike Kap95p, Kap104p bound directly to transport substrate,
without an adaptor. Like Kap95p, Kap104p bound directly to a
subset of peptide repeat containing
nucleoporins. Most importantly,
a mutant Kap104p was rapidly degraded at the nonpermissive temperature
resulting in concomitant failure to import
Nab2p but still allowing import of
a protein containing a classical NLS. Hence yeast Kap104p is
a signal recognition and docking factor for at least Nab2p,
whose NLS still has to be identified. Nab2p or Nab4p do not
contain a region of close similarity
to the previously mapped M9 nuclear
localization sequence of the abundant
human nuclearmRNA binding protein A1
(21). A human homolog of
Kap104p, termed transportin, has recently been described and
shown to be necessary for the nuclear
import of
an M9-carrying reporter protein (22).
2.
2
cDNA.
2
(Kap104p) was obtained from Genome Systems (St. Louis). This clone
coded for the C-terminal region of
karyopherin2.
To obtain the full-length coding sequence, a human liver 5 -Stretch
Plus cDNA library (CLONTECH) was screened according to the manufacturer's
instructions using a single-stranded antisense oligonucleotide
(corresponding to bases 105-155 of
the Expressed Sequence Tag database) that was labeled with [
-Stretch
Plus cDNA library (CLONTECH) was screened according to the manufacturer's
instructions using a single-stranded antisense oligonucleotide
(corresponding to bases 105-155 of
the Expressed Sequence Tag database) that was labeled with [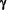 -32P]-ATP
by polynucleotide kinase (23). Three partial overlapping
clones were isolated and cloned in pBlue Script II SK (Stratagene).
The full-length coding sequence for karyopherin2
was determined from these overlapping clones and cDNA clone
224297 and has been deposited in the GenBank database (accession
no. U72069
[GenBank]).
-32P]-ATP
by polynucleotide kinase (23). Three partial overlapping
clones were isolated and cloned in pBlue Script II SK (Stratagene).
The full-length coding sequence for karyopherin2
was determined from these overlapping clones and cDNA clone
224297 and has been deposited in the GenBank database (accession
no. U72069
[GenBank]).
Expression and Purification of Recombinant
Proteins.
2,
a fragment of the clone 224297 was
generated by digestion with BstEII and NotI. This
fragment was ligated between the BstEII and
NotI sites
of pBluescript II SK containing
the longest clone previously isolated from the human liver cDNA
library. To obtain a cDNA coding for a glutathione S-transferase
(GST)-karyopherin2
fusion protein, the full-length karyopherin2
cDNA was amplified by the PCR using the primers 5CACCTCAGGCCCCGGGCCAAGAAGGAG3
and 5GGGACTGCAGCTCGAGTGTATTAGAATAAAA3
(introducing an XmaI and an XhoI site at the 5
and 3 ends, respectively),
and then subcloned in frame in pGEX-4T-3 (Pharmacia). The GST-karyopherin2
fusion protein was purified from Escherichia coli BL21/LysS
by binding to glutathione Sepharose 4B beads (Pharmacia). The
recombinant protein was recovered from the beads by cleavage with
thrombin (Sigma) followed by inactivation of
thrombin by hirudin (Sigma).
-AAAGTCTCTCTTCACCCCCCGGGTCAAGTCTAA-3
and 5-CTCCTGCTAAGCTTTGTTCTCGAGTTAAAATCT-3
introducing an
XmaI and a XhoI site at the 5
and 3 ends, respectively. The PCR
product was subcloned in frame into the
XmaI/XhoI.
sites of pGEX-4T-3. The
GST-A1 fusion protein expressed in E. coli BL21/LysS was purified
by binding to glutathione beads and recovered by elution with
10 mM reduced glutathione. The GST-A1 fusion protein was labeled
with fluorescein isothiocyanate (24).
2,
human wild-type Ran, mutant Ran, p10, and Nup98 (residues 43-824) were
prepared as described (9, 10,
25-28).
Recombinant GST-karyopherin1
was prepared as described (6).
Synthetic Peptides.
Solution Binding Assay.
2
was added alone or together with 25× or 100× molar excess of
the A1 NLS or simian virus 40 NLS synthetic peptides. Recombinant
Nup98 (5 ľg) immobilized on 10 ľl of
Affigel beads (Bio-Rad) was incubated with 1 ľg of
GST-1 or 1 ľg of2
or with 1 ľg of GST-1
and 10× molar excess of2
or with 1 ľg of2
and 10× molar excess of GST-1.
Overlay Blot Assay.
2
(1 ľg/ľl) or 1 (1 ľg/ľl)
previously metabolically labeled with 35S-Express
Protein Labeling Mix (NEN). The blot was washed three × 10 min
in the same buffer and 3 min in transport buffer, then dried
and exposed for autoradiography.
Nuclear Import
Assay.
2;
0.5 ľg ofkaryopherin1;
4 ľg of Ran (wild type or mutant);
60 ng of p10; 0.4 ľg of
NLS-human serum albumin; 1 ľg of GST-A1;
and 4 ľg of wheat germ agglutinin.
2
(Fig. 1). Its calculated molecular mass is
101,321 daltons. Over its entire sequence, human 2
is 34% identical and 50% similar to yeast Kap104p (Fig. 1).
Human 2 is 17% identical
and 30% similar to human 1
(Fig. 1). While this work was in progress, Pollard
et al. (22) reported similar data and proposed
the term transportin for karyopherin2.
The two sequences were identical, except for an isoleucine at
position 217 in transportin that is substituted with a threonine
in karyopherin2. 
Fig. 1. Comparison of amino
acid sequences of human
karyopherin2,
yeast karyopherin2
(Kap104p), and human
karyopherin1.
Sequences were aligned using the CLUSTALW
v.1.6 program and analyzed with BOXSHADE.
Identical amino acids are indicated by black boxes and similar
amino acids by gray boxes.
[View
Larger Version of this Image (110K
GIF file)]
2
and expressed it in
E. coli as a GST fusion protein with a thrombin
cleavage site between the GST and the 2
moiety of the fusion protein.
Thrombin cleavage generated a full-length recombinant 2
(Fig. 2A). Recombinant 2
bound to an immobilized fusion protein consisting of
GST and the A1 protein (Fig.
2B, lane 1). The
NLS of A1 has been sublocalized
to its C-terminal region and termed M9 (21). A
synthetic peptide representing this region inhibited binding of
recombinant 2 to A1 (Fig.
2B, lane 2) whereas a classical NLS
peptide had no effect (Fig. 2B, lane 3). We conclude
that karyopherin2
is a signal recognition factor that specifically recognizes
the A1 type NLS, but not the classical NLS, confirming previous
data (22). 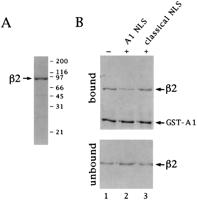
Fig. 2. Karyopherin2
binds to a fusion protein (GST-A1) containing the nuclear
mRNA binding protein A1 via a specific
sequence. (A) Purified recombinant karyopherin2
analyzed by SDS/PAGE and visualized by Coomassie blue staining.
(B) Immobilized GST-A1 was incubated with karyopherin2
alone (lane 1) or with a 25× molar excess of
a synthetic peptide representing the NLS of
the A1 protein (lane 2) or with a 100× molar excess of
the classical NLS peptide (lane 3). Bound and unbound fractions
were analyzed by SDS/PAGE and Coomassie blue staining.
[View
Larger Version of this Image (38K GIF
file)]
2
is also a docking factor as it binds to a subset of
nucleoporins. This was shown by using SDS/PAGE-separated nuclear
envelope proteins that were transferred to nitrocellulose and
were probed in overlay blots with metabolically labeled [35S]karyopherin2
(Fig.
3A, lane 1). For comparison, the same blot
was also probed with metabolically labeled [35S]karyopherin1
(lane 2) that was previously shown in this assay to bind to
a subset of peptide repeat containing
nucleoporins (10). Like karyopherin1,
karyopherin2
bound to several proteins that comigrated with known peptide
repeat-containing nucleoporins.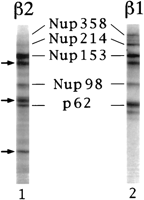
Fig. 3. Karyopherin2
binds to peptide repeat-containing nucleoporins. Proteins from purified
rat liver
nuclear envelopes were separated
by SDS/PAGE, transferred to nitrocellulose, and incubated with
35S-labeled karyopherin2
(lane 1) or 35S-labeled karyopherin1
(lane 2). The arrows indicate positions of
unidentified bands that interact with
karyopherin2.
[View
Larger Version of this Image (35K GIF
file)]
1
has previously been mapped to the peptide repeat-containing region of
the nucleoporin Nup98 (26). To determine
whether 2 also bound to
repeat regions we used E. coli lysates that contained
recombinant regions of Nup98
(26) and probed them in an overlay blot with 35S-labeled 2.
As previously reported for 1, 2
bound to the N-terminal fragment of
Nup98 that contains the peptide repeat region, but not to the
repeat-lacking C-terminal region of
Nup98 (data not shown). When Nup98 was purified and immobilized,
it bound GST-1 or 2
in a solution binding assay (Fig. 4B, lanes 1
and 2). Interestingly, both 1
and 2 competed with each
other for binding (lanes 3 and 4). Together these data suggest
that 1 and 2
bound to similar or overlapping sites in the peptide repeat
region of Nup98. 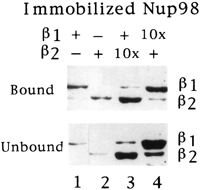
Fig. 4. Karyopherin2
and 1 compete for binding
to Nup98. Immobilized Nup98 was incubated with GST-1
(lane 1), 2 (lane 2), GST-1
and 10× molar excess of2
(lane 3), or 2 and 10× molar
excess of GST-1
(lane 4). Bound and unbound fractions were analyzed by SDS/PAGE
and Coomassie blue staining.
[View
Larger Version of this Image (49K GIF
file)]
2.
Nuclear rim staining was observed
only in the presence of2
(Fig. 5 A, 1 and 2) indicating that 2
is required for docking of GST-A1
at nucleoporins. In contrast, there was no 2-mediated
docking of fluorescently
labeled, classical NLS-containing substrate [NLS-human serum
albumin (24)], either in the absence or presence of
karyopherin
(data not shown). Hence,
karyopherin2
is both a signal recognition and docking factor that specifically
recognizes A1's NLS and docks A1 to repeat containing nucleoporins
without requiring an energy generating system. 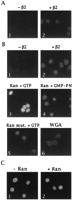
Fig. 5. GST-A1 import into
the nucleus requires
karyopherin2
and Ran. (A) Digitonin-permeabilized HeLa cells were incubated at
4°C with fluorescently labeled GST-A1, in the presence or absence
of karyopherin2
(2 ľg/assay) as indicated. (B) Permeabilized cells were
incubated at 20°C, with fluorescently labeled GST-A1 in the
absence (panel 1) or in the presence of
karyopherin2
(2 ľg/assay)
[View
Larger Version of this Image (30K GIF
file)]
2,
and in the absence or presence of recombinant
Ran. In the absence
of both 2
and Ran there was no readily detectable import
of GST-A1 into nuclei (Fig. 5B1).
Together with the docking data this result indicates that the
digitonin-permeabilized cells contained either little endogenous 2
or that endogenous A1-type NLS substrate was inefficiently displaced
by GST-A1. In the presence of
low concentrations of2
(0.5 ľg/assay) there was import (Fig.
5C1) which was not noticeably stimulated
by exogenously added Ran (Fig. 5C2).
The imported GST-A1 was distributed
throughout the nucleoplasm but apparently was excluded from
nucleoli. These negatively stained nucleoli served as a useful
criterion for 2-mediated
GST-A1 import. At higher concentrations
of2
(2 ľg/assay) and in the absence of
added Ran there was primarily docking at the nuclear
rim and little, if any, import based
on the absence of negatively
stained nucleoli (Fig.
5B2), whereas in the presence
of Ran there was a striking stimulation
of import
with the appearance of negatively
stained nucleoli (Fig. 5B3). Similar results were
obtained at still higher concentrations of2
(4 ľg/assay)(data not shown). These data indicate that at low
levels of added 2
(0.5 ľg/assay) the endogenous Ran of
the digitonin-permeabilized cells may suffice for maximal import,
whereas exogenously added Ran is required to achieve maximal
levels of import
at higher concentrations of
exogenously added 2. Import
was inhibited in the presence of
the nonhydrolyzable guanylyl imidodiphosphate (Fig. 5B4)
or by the exogenous addition of mutant
Ran that is unable to hydrolyze GTP (Fig. 5B5).
Wheat germ agglutinin inhibited import
but still allowed 2-mediated
docking (Fig. 5B6).
1
and 2 competed with each
other for binding to the repeat containing domain of
Nup98 (and likely also to those of
other repeat-containing nucleoporins), suggested that 1
and2 also may compete with each
other in nuclear import
by competing for common or overlapping nucleoporin docking sites.
Indeed, import of
NLS-human serum albumin, mediated by karyopherin2, 1,
Ran, and p10 (10) (Fig. 6A1)
was inhibited by2
(Fig. 6A2) and vice versa, import
of GST-A1, mediated by2
and and Ran (Fig. 6B1) was inhibited by 1
(Fig. 6B2). Hence, the distinct/1-
and 2-mediated pathways
of nuclear
import for their substrates
appear to at least partially merge at the level of
docking to nucleoporins. 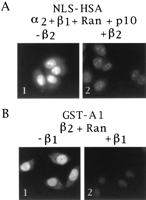
Fig. 6. Karyopherin1
and karyopherin2
compete for
nuclear import.
(A) Digitonin-permeabilized HeLa cells were incubated at 20°C
with fluorescently labeled NLS-human serum albumin,
karyopherin2,
karyopherin1,
Ran, and p10 in the absence (panel 1) or in the presence of
10× molar excess of karyopherin2
(panel 2). (B) Digitonin-permeabilized HeLa cells were
incubated with fluorescently labeled GST-A1, karyopherin2,
and Ran in the absence (panel 1) or in the presence of
10× molar excess ofkaryopherin1
(panel 2). (panels 2-6). Wild-type Ran (panels 3, 4, and 6)
or a GTPase-deficient mutant Ran (panel 5) were added. Wheat
germ agglutinin (WGA) was added in panel 6. (C) Permeabilized
cells were incubated at 20°C with fluorescently labeled GST-A1
and karyopherin2
(0.5 ľg/assay) in the presence or absence of
Ran as indicated.
[View
Larger Version of this Image (56K GIF
file)]
1, 2,3,
and 4 for Kap95p, Kap104p,
Pse1p, and Kap123p, respectively. It seems likely that each
of the karyopherins
recognizes its own type of
NLS. Should this be the case then, as a further simplification of
nomenclature, the corresponding NLSs might be termed NLS-1, NLS-2,
NLS-3, and NLS-4, respectively. It appears that karyopherin2, 3,
and 4 bind directly their
cognate NLSs (ref. 19; M. P. Rout, G.B.,
and J. D. Aitchison, unpublished data). In contrast, karyopherin1
uses
karyopherin
as an adaptor for binding to the NLS-1 substrate. In yeast there
is only one karyopherin,
whereas in mammalian cells there are at least two
karyopherins, which may
have distinct, but overlapping, substrate specificities (10).
All karyopherins
bind directly to similar (but not always identical) repeat nucleoporins.
Therefore, all three (or four) presently known import
pathways appear to merge at the level of
docking of the various
karyopherins to similar or overlapping
repeat domains of some, but not
necessarily identical, repeat nucleoporins. These repeat nucleoporins
are distinct components of
the fibers emanating from the nucleoplasmic and the cytoplasmic side
of the nuclear
pore complex (reviewed in ref. 30). Hence the
karyopherins function to concentrate
transport substrate at multiple docking sites of
the nuclear pore complex fibers. This
fibrous zone would serve as an atrium to the central opening of
the nuclear pore complex. Cytoplasmic
proteins lacking NLSs for karyopherin-mediated
docking (or lacking sites for direct docking) to repeat nucleoporins
might be sterically excluded from this atrium and therefore
would be prevented from entering the nucleus. Steric exclusion
from the atrium would be more efficient for large cytoplasmic
proteins and less efficient for small cytoplasmic proteins,
which therefore may enter the nucleus without an NLS or without
a docking site to repeat nucleoporins.
2.
Similar to yeast 2 (19),
this mammalian2 bound directly
to NLS-2 substrate (the mRNA-binding
protein A1) and to nucleoporins. In solution binding assays,
binding to the NLS-2 substrate could be competed for by NLS-2
peptide, but not by NLS-1 peptide, indicating that 2
binds specifically to NLS-2, but does not bind to NLS-1 (see
also ref. 22). In overlay blots, mammalian 2
apparently bound to some of the same
repeat nucleoporins, which previously had been shown to bind
to 1 (10),
though there were distinct differences in the binding affinity.
For example, although binding to the nucleoplasmically exposed
nucleoporins Nup98 and Nup153 was similar for 2
and 1, binding to
Nup358 and Nup 214, cytoplasmically exposed nucleoporins, was much
weaker for 2 than it was
for 1. The significance
of these affinity differences
for both
nuclear import
and export remains to be elucidated. For one nucleoporin, Nup98,
we have localized2 binding to
the N-terminal repeat motif containing region of
Nup98. It is likely that binding to other repeat motif-containing
nucleoporins is also to their repeat regions, although this remains
to be shown.
2 was able
to dock the fluorescently labeled GST-A1 protein at the nuclear
rim of digitonin-permeabilized cells
at 0°C. At 20°C and in the presence of
GTP and an energy-generating system, fluorescently labeled GST-A1
was not imported into nuclei unless 2
was present in the import reaction.
At low concentration of2,
exogenously added Ran did not stimulate import.
However, at higher concentrations of2
there was primarily docking and, based on the absence of
negatively stained nucleoli, virtually no import.
Strikingly, the addition of Ran greatly
stimulated import and diminished
docking. These data suggested that Ran is required for2-mediated
import and that at lower concentration
of2
the endogenous Ran is sufficient for import
but that at higher concentration of2
exogenously added Ran is required for maximal
import.
It is presently not clear why the import
that is likely to be mediated by endogenous Ran appears to be
inhibited at high concentrations of2.
2-mediated
import as nonhydrolyzable guanylyl
imidodiphosphate or the addition of
a Ran mutant that is unable to hydrolyze GTP inhibited import.
The exact function of Ran
and GTP hydrolyisis in 2-mediated
import remains to be elucidated.
Unlike 1, which binds Ran-GTP
with high affinity in a solution binding assay (16,
27), high affinity binding of
Ran-GTP to 2 could not be
detected (data not shown). Moreover, Ran-GTP does not appear
to dissociate 2 bound to
immobilized GST-A1 and p10 does not stimulate2-mediated
import into nuclei of
digitonin-permeabilized cells as it does in the case of/1-mediated
import of
NLS-1 protein (data not shown). One possibility is that Ran
stimulation of2-mediated
import is indirect and is due to
Ran-mediated clearance
of endogenous /1
and of3
[which also binds Ran-GTP (31)] from nucleoporin
docking sites. However the finding that preincubation of
digitonin-permeabilized cells with Ran-GTP did not abolish Ran
stimulation of GST-A1 import
at the higher concentration of2
(2 ľg/assay) (data not shown) argues against this possibility.
2
functions as a signal recognition and docking factor that binds directly
to an NLS-2 containing substrate and to peptide repeat-containing
nucleoporins. Karyopherin2-
mediated docking of NLS-2 substrate
at the nuclear rim of
digitonin-permeabilized cells occurs on ice and does not require
energy. Import is stimulated
by Ran and GTP hydrolysis. Not all nuclearmRNA
binding proteins are imported via the
NLS-2 pathway (32). Moreover, proteins other
than nuclear mRNA
binding proteins may be
imported
via the NLS-2 pathway.
FOOTNOTES
ACKNOWLEDGEMENTS
We thank John Aitchison, Lucy Pemberton, and Michael Rout for advice,
helpful discussions, and critical reading of
the manuscript; Philip Bernstein for providing us with purified
mutant Ran; Karsten Weis and Angus Lamond for the karyopherin2/hSRP1a-expressing
vector, and Stephen Adam for the karyopherin1/p97
expressing vector.
ABBREVIATIONS
NLS, nuclear localization sequence;
GST, glutathione S-transferase.
REFERENCES
| 1. | Adam, E. J. & Adam, S. A. (1994) J. Cell Biol.125, 547-555[Abstract] |
| 2. | Görlich, D., Prehn, S., Laskey, R. A. & Hartmann, E. (1994) Cell79, 767-778[Medline] |
| 3. | Radu, A., Blobel, G. & Moore, M. S. (1995) Proc. Natl. Acad. Sci. USA92, 1769-1773[Medline] |
| 4. | Moroianu, J., Blobel, G. & Radu, A. (1995) Proc.
Natl. Acad. Sci. USA2, 2008-2011 |
| 5. | Görlich, D., Kostka, S., Kraft, R., Dingwall, C., Laskey, R. A., Hartmann, E. & Prehn, S. (1995) Curr. Biol.5, 383-392[Medline] |
| 6. | Chi, N. C., Adam, E. J. & Adam, S. A. (1995) J. Cell Biol.130, 265-274[Abstract] |
| 7. | Imamoto, N., Tachibana, T., Matsubae, M. & Yoneda, Y. (1995) J. Biol. Chem.270, 8559-8565[Abstract/Full Text] |
| 8. | Imamoto, N., Shimamoto, T., Takao, T., Tachibana, T., Kose, S., Matsubae, M., Sekimoto, Y. & Yoneda, Y. (1995) EMBO J.14, 3617-3626[Medline] |
| 9. | Weis, K., Mattaj, I. W. & Lamond, A. I. (1995) Science268, 1049-1053[Medline] |
| 10. | Moroianu, J., Hijikata, M., Blobel, G. & Radu, A. (1995) Proc. Natl. Acad. Sci. USA92, 6532-6536[Medline] |
| 11. | Moore, M. S. & Blobel, G. (1993) Nature (London)365, 661-663[Medline] |
| 12. | Melchior, F., Paschal, B., Evans, J. & Gerace, L. (1993) J. Cell Biol.123, 1649-1659[Abstract] |
| 13. | Moore, M. S. & Blobel, G. (1994) Proc. Natl. Acad. Sci. USA91, 10212-10216[Medline] |
| 14. | Paschal, B. M. & Gerace, L. (1995) J. Cell Biol.129, 925-937[Abstract] |
| 15. | Görlich, D., Vogel, F., Mills, A. D., Hartmann, E. & Laskey, R. A. (1995) Nature (London)377, 246-248[Medline] |
| 16. | Rexach, M. & Blobel, G. (1995) Cell83, 683-692[Medline] |
| 17. | Nehrbass, U. & Blobel, G. (1996) Science272, 120-122[Abstract] |
| 18. | Enenkel, C., Blobel, G. & Rexach, M. (1995) J. Biol. Chem.270, 16499-16502[Abstract/Full Text] |
| 19. | Aitchison, J. D., Blobel, G. & Rout, M. P. (1996) Science274, 624-627[Abstract/Full Text] |
| 20. | Chow, T. Y., Ash, J. J., Dignard, D. & Thomas, D. Y. (1992) J. Cell Sci.101, 709-719[Medline] |
| 21. | Siomi, H. & Dreyfuss, G. (1995) J. Cell Biol.129, 551-559[Abstract] |
| 22. | Pollard, V., Michael, M. W., Nakielny, S., Siomi, M. C., Wang, F. & Dreyfuss, G. (1996) Cell86, 985-994[Medline] |
| 23. | Sambrook, J., Fritsch, E. F. & Maniatis, T. (1989) Molecular Cloning: A Laboratory Manual (Cold Spring Harbor Lab. Press, Plainview, NY), 2nd Ed. |
| 24. | Moore, M. S. & Blobel, G. (1992) Cell69, 939-950[Medline] |
| 25. | Coutavas, E., Ren, M., Oppenheim, J. D., D'Eustachio, P. & Rush, M. G. (1993) Nature (London)366, 585-587[Medline] |
| 26. | Ren, M., Drivas, G., D'Eustachio, P. & Rush, M. G. (1993) J. Cell Biol.120, 313-323[Abstract] |
| 27. | Floer, M. & Blobel, G. (1995) J. Biol. Chem.271, 5313-5316[Abstract/Full Text] |
| 28. | Radu, A., Moore, M. S. & Blobel, G. (1995) Cell81, 215-222[Medline] |
| 29. | Matunis, M. J., Coutavas, E. & Blobel, G. (1996) J. Cell Biol.135, 1457-1470[Abstract] |
| 30. | Rout, M. P. & Wente, S. R. (1994) Trends Cell Biol.4, 357-365 |
| 31. | Yaseen, N. R. & Blobel, G. (1997) Proc. Natl. Acad.
Sci. USA94, 4489-4494 |
| 32. | Pi?ol-Roma, S. & Dreyfuss, G. (1993) Trends Cell Biol.3, 151-155 |
|
|
This article has been cited by other articles: